

-
Paul Lindahl
- Professor, Chemistry, Biochemistry and Biophysics
- Focus Area: bioinorganic, biophysical, bioanalytical, metal ions, trafficking, math modeling, Mössbauer spectroscopy, chromatography, ICP-MS, mitochondria, labile metal pools, ironomics, Saccharomyces cerevisiae, Jurkat cells, hemochromatosis, mice
- Office:
- CHEM 1129
- Email:
- [email protected]
- Phone:
- 979-845-0956
Education
- Undergraduate Education
- B.A. from North Park College, Chicago IL in 1979
- Graduate Education
- Ph.D. from the Massachusetts Institute of Technology in 1985
- Postdoc at the University of Minnesota, from 1985-1988
Areas of Expertise
- Bioinorganic
- Biophysical
- Bioanalytical
- metal ions
- trafficking
- math modeling
- Mössbauer spectroscopy
- Chromatography
- CP-MS, mitochondria
- labile metal pools
- ironomics
- Saccharomyces cerevisiae
- Jurkat cells
- Hemochromatosis
- mice
Professional Summary
Transition metals play essential roles in cells; they have outstanding catalytic properties and are often found at the active sites of enzymes. These metals are also dangerous to cells, as they can generate reactive oxygen species that are associated with diseases and aging. The Lindahl lab is interested in metal ion trafficking and homeostasis in bacteria, yeast, human cells, and mice. We employ Mössbauer spectroscopy to probe the iron content of cells; this powerful biophysical method can “see” the distribution and speciation of iron in a cell – the ironome. Bioanalytical chemistry is used to identify and characterize “labile metal pools” – nonproteinaceous metal complexes that are used in trafficking, signaling, and regulation. Such complexes are detected using a liquid chromatography system interfaced to an inductively-coupled plasma mass spectrometer. We use these methods to study an iron-overload disease in genetically modified mice. Finally, we are developing differential equations–based mathematical models of iron trafficking kinetics.
Biophysics of iron in biology

High-field Mössbauer spectrometer
Iron is a redox-active transition metal with superior catalytic properties that are essential for processes involving redox reactions, oxygen-activation, bioenergetics, metabolism, and genomic stability. Mössbauer spectroscopy involves nuclear spin transitions similar to NMR. It is the most powerful spectroscopy to study iron and we use it to probe the iron content of cells. Mössbauer can differentiate hemes, iron-sulfur clusters, nonheme iron, di-iron oxo centers. It can also assess oxidation states and electronic spin states. We use this tool to understand better the trafficking and regulation of iron in cells and isolated organelles such as mitochondria. We use various genetic strains of yeast, which are grown under different conditions to probe the mechanism of iron trafficking and regulation.
J. D. Wofford, N. Bolaja, N. Dziuba, F. W. Outten, P. A. Lindahl, Evidence that a respiratory shield in Escherichia coli protects a low molecular mass Fe(II) pool from O₂-dependent oxidation. J. Biol. Chem. 294, 50–62 (2019).
Labile metal pools

LC-ICP-MS system
Cells contain low molecular weight, nonproteinaceous metal complexes (metal ions bound to metabolite ligands using coordinate bonds) that are used in metal ion trafficking, signaling/sensing, and regulation. Despite decades of study, the chemical identity and precise roles of these complexes in cellular metabolism are unknown. We have built a novel liquid chromatography system installed in an anaerobic glove box that is interfaced to an inductively-coupled plasma mass spectrometer. We use this LC-ICP-MS system to detect these complexes, and then use electrospray ionization mass spectrometry of fractions off the column to identify them. We use the same approach to study low-mass copper, zinc, and manganese complexes.
T. Q. Nguyen, J. E. Kim, H. N. Brawley, P. A. Lindahl, Chromatographic detection of low-molecular-mass metal complexes in the cytosol of Saccharomyces cerevisiae. Metallomics (2020).
Mathematical modeling
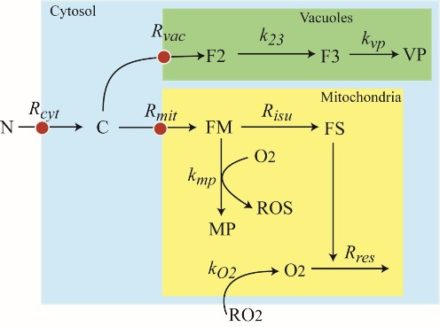
Model developed by J. D. Wofford and P. A. Lindahl.
See BMC Syst. Biol. 13 (2019).
Budding yeast S. cerevisiae contains about 100 proteins in which iron centers (hemes, iron-sulfur clusters, di-iron-oxo centers, etc.) are installed. We have cataloged these proteins and know their location in the cell (cytosol, mitochondria, nucleus, endoplasmic reticulum, and vacuoles). We also know their approximate concentration. In this project, we are developing ordinary differential equations–based kinetic models to predict the concentration of each iron species in the cell under steady-state and time-dependent (dynamical) conditions. Because all iron-based reactions in the cells will be included in the model, we expect that it will show a more sophisticated ability to understand the observed phenotypes of various iron-related genetic mutants. The models will be optimized against experimental results and will be used to predict the outcomes of new experiments and be tested by them. Simpler models that are limited to mitochondrial iron-based reactions have been used to understand the progress of the disease Friedreich’s ataxia, according to the mechanism shown in the figure.
J. D. Wofford and P. A. Lindahl, A mathematical model of iron import and trafficking in wild-type and Mrs3/4ΔΔ yeast cells. BMC Syst. Biol. 13 (2019).
Mouse studies
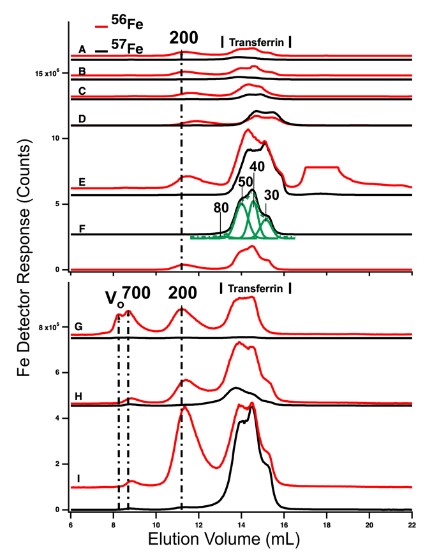
LC-ICP-MS of iron complexes in blood.
See Metallomics 11, 1900–1911 (2019).
Hemochromatosis is a disease affecting 1 in 300 individuals in which too much nutrient iron enters the blood. Blood contains an iron-buffering protein called transferrin, which in healthy individuals “soaks up” excess nutrient iron. In patients with hemochromatosis, transferrin becomes saturated and extra iron enters the blood in a nonproteinaceous form called “non-transferrin-bound iron” or NTBI. NTBI is absorbed by the liver and eventually the heart. Its chemical composition remains unknown despite decades of research. In this project, we are comparing the blood plasma from mice that are healthy with those with hemochromatosis using our LC-ICP-MS system. We are also investigating the form of iron that overloads organs using Mössbauer spectroscopy. The goal is to identify NTBI and better understand its properties in the blood and how it affects mouse organs. Mice are dissected in an anaerobic glove box to avoid iron oxidation. We have detected several low-mass iron complexes but none with all of the characteristics of NTBI.
N. Dziuba, J. Hardy, P. A. Lindahl, Low-molecular-mass iron complexes in blood plasma of iron-deficient pigs do not originate directly from nutrient iron. Metallomics 11, 1900–1911 (2019).
All Publications
- View publications on Web of Science