

-
Juan Dong
- Sadie Hatfield Professor
- Focus Area: Molecular Genetics, Cell Signaling, Cell Biology, Plant Development
- Office:
- 302A
- Email:
- [email protected]
- Phone:
- 979-314-8279
Education
- Undergraduate Education
- Shandong Normal University, China (1997)
- Graduate Education
- M.S. Institute of Botany, Chinese Academy of Sciences (2000)
- Ph.D. University of California, Riverside (2005)
- Postdoc. Stanford University (2011)
Areas of Expertise
Molecular Genetics, Cell Signaling, Cell Biology, Plant Development
Professional Summary
Cell polarity, in both animals and plants, is of paramount importance for developmental and physiological processes. The establishment and maintenance of cell polarity is required for asymmetric cell division (ACD) and indispensable for multi-cellular organisms to generate cellular diversity. Through ACD, a single mother cell can produce daughter cells with distinctive identities in developmental differentiation. Our research focuses on the mechanisms by which cell polarity is initiated and maintained in the stomatal lineage cells in Arabidopsis and the mechanisms by which differential cell fates are specified in plant ACD. The identification of the plant-specific protein BASL (Breaking of Asymmetry in the Stomatal Lineage) provided strong evidence that plant cells have the capability to polarize non-transmembrane proteins and utilize such polarized protein distribution to regulate many cellular events during asymmetric cell division (Figure 1).

Figure 1. BASL localization and stomatal asymmetric cell fate
By using BASL as an anchor for screens to isolate genetic and physical interactors and by using the features of the protein itself as a probe for cell’s ability to correctly establish polar cortical localization, our lab is in the process of building a model for plant cell polarization and its regulation in ACD. We are also interested in understanding the molecular mechanisms underlying how cell polarity orients cell division plant and how differential daughter cell fates are specified by the cortical polarity complex. Our work focuses on the identification of additional polarized proteins and of mutants that display specific subsets of polarity defects. At a larger extent, some of our work also include the characterization of novel regulators in stomatal development in Arabidopsis. Through collaborative effort, we also expand our research interest towards understanding the mechanical and physiological features of functional stomatal guard cells in plants. During 2021-2022, we made major contributions below a few important areas in the cell polarity and asymmetric cell division field.
Membrane trafficking for polarity initiation and establishment in plant cells
The molecular mechanisms for protein polarization in plants have been extensively studied in two systems: 1) vesicular trafficking-based polarization of PIN auxin efflux carriers (membrane embedded proteins) and 2) cytoskeleton-dependent and –independent positive feedback loop-based ROP polarization (small Rho-like GTPases from plants). Polar trafficking of BASL, a non-membrane novel protein, has not been successfully connected to both pathways and might represent an unknown mechanism. Fluorescence Recovery After Photobleaching (FRAP) was performed on GFP-BASL and the recovery curves suggested that BASL dynamics is more comparable to the membrane-embedded PIN proteins, hinting the possible regulation of membrane trafficking in BASL polarization. Indeed, through the identification of physical interactors of BASL, we recently established that four members of the PRAF protein family interact with BASL and are required for the establishment of the polarity site (Wang et al., 2022 Nat. Commun) (Figure 2). The PRAF proteins are plant specific but contain phospholipid-binding domains and co-localize with small GTPases, both of which are conserved across the kingdoms in regulating membrane trafficking and delivery.

Figure 2. A working model for PRAF-mediated trafficking in establishing BASL polarity. In wild-type plants, the polarization of BASL protein (green) in the stomatal lineage cell requires the physical partner, four PRAF proteins (orange), as well as the Golgi-localized Arf GEF GNOM (blue). The PRAF8 proteins are predominantly localized to the plasma membrane, where they may polarize together with BASL. The PRAF8 proteins may also partially associate with the Golgi, TGN/EE and a subset of endosomes/vesicles decorated by RabC1 and RabF2b. Furthermore, the PRAF proteins physically interact with GNOM, possibly leading to the association of GNOM to the RabC1- and RabE1d-decorated membrane structures.
BASL scaffolds the BSL phosphatases to enable the progression of stomatal asymmetric cell division
Our previous work showed that BASL is phosphorylated and activated by MAPK 3 and 6 (MPK3/6) and becomes polarized to the cell cortex, where it recruits the MAPKK Kinase YODA and MPK3/6 to inhibit stomatal differentiation in one of the two daughter cells. Recent work from other labs showed that, prior to a stomatal ACD, the polarity complex employs POLAR to recruit the GSK3-like kinase BIN2 that releases the suppression of YODA on stomatal differentiation, so that stomatal ACD is promoted. Therefore, the stomatal polarity complex by scaffolding different signaling molecules could promote the division potential before an ACD and suppress the division potential after an ACD. However, how the transition of these two seemingly opposing procedures can be achieved by the same polarity complex remained a major challenge towards understanding stomatal ACD. Here, by using immunoprecipitation combined with mass spectrometry (IP-MS), we identify a family of protein Ser/Thr BSL phosphatases, as BASL partners (Guo, 2021 Nat Plants). Genetic analysis places BSL upstream of the YDA MAP kinase cascade and downstream of the plasma membrane receptors. In addition, the founding member BSL1 colocalizes with BASL in a polarized manner at the cell periphery. Interestingly, the recruitment of the BSL phosphatases in the polarity module confers a negative role to BIN2 complex but a positive role to the YDA MAPK module (Figure 3). Thus, our study reveals a crucial function of the BSL phosphatases in bridging the two opposing protein functional modules to control the balance of cell-division potential and cell-fate determination in plant ACDs. This work also represents the initiation of collaboration with the Nickels lab at the Waksman Institute.
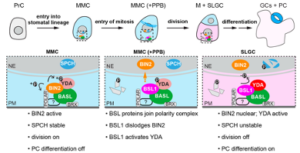
Figure 3. A working model for BSL phosphatases to function in stomatal ACD. The BSL proteins function as the spatiotemporal molecular switch enabling the coordination of cell division and cell-fate differentiation in stomatal ACD. BSL1 join the polarity complex in the ACD mother cells that are committed to cell division. Association of BSL1 with the polarity complex dislodges BIN2 from the cell membrane to prevent subsequent rounds of cell division and activates the YDA MAPK signaling cascade to promote cell-fate differentiation. BIN2, GSK3-like kinase; YODA, MAPK Kinase Kinase; SPCH, bHLH transcription factor; BASL and
Signaling dichotomy of the BSL phosphatases in controlling stomatal fate determination
MAPK signaling modules play crucial roles in regulating numerous biological processes in all eukaryotic cells. How MAPK signaling specificity and strength are tightly controlled remains a major challenging question. In Arabidopsis stomatal development, the MAPKK Kinase YODA (YDA) functions at the cell periphery to inhibit stomatal production by activating MAPK 3 and 6 (MPK3/6) that directly phosphorylate stomatal fate-determining transcription factors for degradation in the nucleus. Recently, we demonstrated that BSL1, one of the four BSL protein phosphatases, localizes to the cell cortex to activate YDA, elevating MPK3/6 activity to suppress stomatal formation. In this study, we showed that at the plasma membrane, all four members of BSL proteins contribute to the YDA activation. However, in the nucleus, specific BSL members (BSL2, BSL3, and BSU1) directly deactivate MPK6 to counteract the linear MAPK pathway, thereby promoting stomatal formation (Figure 4). Thus, the pivotal MAPK signaling in stomatal fate determination is spatially modulated by a signaling dichotomy of the BSL protein phosphatases in Arabidopsis, providing a prominent example of how MAPK activities are integrated and specified by signaling compartmentalization at the subcellular level (Guo et al., 2022, Nat Commun).
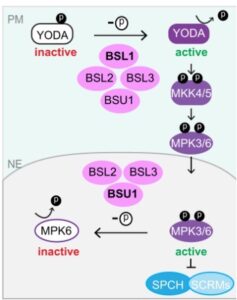
Figure 4. A working model: BSL phosphatases-based signaling dichotomy, through spatial compartmentalization of the regulation on distinct components of the linear YODA MAPK pathway, controls stomatal development in Arabidopsis. At the cell cortex close to the PM, BSL1 is a predominant regulator, together with the other three BSL phosphatases, activating the MAPKKK YODA to promote MAPK signaling. Activated MPK3/6 molecules phosphorylate the key stomatal fate transcription factors, SPCH and ICE1/SCRMs, for degradation, thereby suppressing stomatal production. In the nucleus, BSU1 plays a primary role, together with BSL2 and BSL3, deactivating MPK3/6, resulting in stabilized SPCH and ICE1/SCRMs, thereby promoting stomatal production.